Structure of active β-arrestin-1 bound to a G-protein-coupled receptor phosphopeptide
By Arun K. Shukla, Aashish Manglik, Andrew C. Kruse, Kunhong Xiao, Rosana I. Reis, Wei-Chou Tseng, Dean P. Staus, Daniel Hilger, Serdar Uysal, Li-Yin Huang, Marcin Paduch, Prachi Tripathi-Shukla, Akiko Koide, Shohei Koide, William I. Weis, Anthony A. Kossiakoff, Brian K. Kobilka & Robert J. Lefkowitz.
Published in Nature 497 (7447) on April 21, 2013: 137-41. Link to publication page. PMID: 23604254. PMCID: PMC3654799.
Core Facility: Synthetic Antigen Binder (SAB) Generation and Crystallography
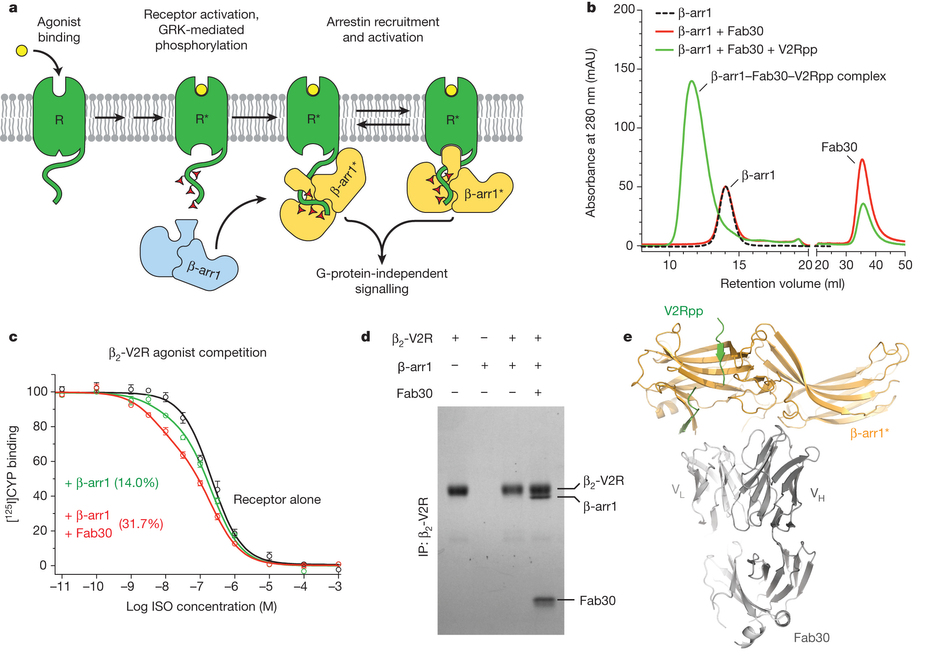
Figure 1: Fab30 specifically recognizes and stabilizes an active state of β-arrestin-1. a, GPCRs are phosphorylated after activation, leading to the binding of arrestins. Interactions between the phosphorylated receptor and β-arrestin-1 lead to β-arrestin-1 activation and the subsequent blockade of G-protein signalling and initiation of β-arrestin-1 signalling pathways. b, Interaction between β-arrestin-1 and Fab30 requires the presence of V2Rpp in a size exclusion assay. c, The formation of a complex between a GPCR and β-arrestin allosterically leads to an enhanced affinity of agonist for the receptor, termed the ‘high agonist affinity state’. Therefore, the fraction of receptor in the high-agonist affinity state reflects the extent of complex formation between receptor and β-arrestin. In a radioligand competition binding assay using 125I-cyanopindolol as the probe and the agonist isoproterenol (Iso) as the competitor, β-arrestin-1 alone shifts a small portion (14%) of receptors into the high agonist affinity state. Fab30 significantly amplifies this effect (31%) (n = 3, P < 0.0001 in F test). d, In a pull-down assay, phosphorylated β2–V2R chimaera shows appreciable binding to β-arrestin-1 only in the presence of Fab30. e, Overall structure of the β-arrestin-1–V2Rpp–Fab30 complex.
Abstract
The functions of G-protein-coupled receptors (GPCRs) are primarily mediated and modulated by three families of proteins: the heterotrimeric G proteins, the G-protein-coupled receptor kinases (GRKs) and the arrestins. G proteins mediate activation of second-messenger-generating enzymes and other effectors, GRKs phosphorylate activated receptors, and arrestins subsequently bind phosphorylated receptors and cause receptor desensitization. Arrestins activated by interaction with phosphorylated receptors can also mediate G-protein-independent signalling by serving as adaptors to link receptors to numerous signalling pathways. Despite their central role in regulation and signalling of GPCRs, a structural understanding of β-arrestin activation and interaction with GPCRs is still lacking. Here we report the crystal structure of β-arrestin-1 (also called arrestin-2) in complex with a fully phosphorylated 29-amino-acid carboxy-terminal peptide derived from the human V2 vasopressin receptor (V2Rpp). This peptide has previously been shown to functionally and conformationally activate β-arrestin-1 (ref. 5). To capture this active conformation, we used a conformationally selective synthetic antibody fragment (Fab30) that recognizes the phosphopeptide-activated state of β-arrestin-1. The structure of the β-arrestin-1-V2Rpp-Fab30 complex shows marked conformational differences in β-arrestin-1 compared to its inactive conformation. These include rotation of the amino- and carboxy-terminal domains relative to each other, and a major reorientation of the ‘lariat loop’ implicated in maintaining the inactive state of β-arrestin-1. These results reveal, at high resolution, a receptor-interacting interface on β-arrestin, and they indicate a potentially general molecular mechanism for activation of these multifunctional signalling and regulatory proteins.
 accession 1G4M; chain A, light blue) and active β-arrestin-1 (gold) were aligned on the N domains. The β-arrestin-1 C terminus is highlighted in dark blue. a, A substantial rotation and translation of the C domain relative to the N domain occurs upon activation. The rotation axis is indicated as a solid black line. b, View of C-domain rotation along the axis. c, N domain of inactive arrestin, highlighting important regions. d, Active β-arrestin-1 in the same orientation, showing V2Rpp in green. Phosphorylated residues are highlighted as sticks. e, The overall structure of inactive β-arrestin-1 (PDB 1G4M; chain A), with loops from all inactive β-arrestin-1 structures superimposed (grey loops). The active conformation of these loops (orange loops) deviates from all inactive structures.")
 displaces the inactive finger loop (light blue), causing it to adopt an extended conformation in the active state (gold). Select charge–charge contacts are shown with dotted lines in b–f. c, In the inactive conformation, the β-arrestin-1 C-terminal β-strand (dark blue) lies along the N domain in the three-element interaction network. d, Upon activation, this strand is displaced by the C terminus of V2Rpp, which engages in extensive charge–charge interactions through phosphorylated residues. e, The polar core of β-arrestin-1 is thought to be a critical stabilizer of the inactive state. f, Upon V2Rpp binding, the C-terminal strand residue Arg 393 is displaced, and its interaction partner D297 undergoes a large movement together with the rest of the lariat loop.")
