A novel mechanism for fine-tuning open-state stability in a voltage-gated potassium channel
By Stephan A. Pless, Ana P. Niciforovic, Jason D. Galpin, John-Jose Nunez, Harley T. Kurata & Christopher A. Ahern.
Published in Nature Communications April 30, 2013;4:1784. PMID: 23653196. PMCID: PMC3644096. Link to publication page.
Core Facility: Membrane Protein Expression/Purification
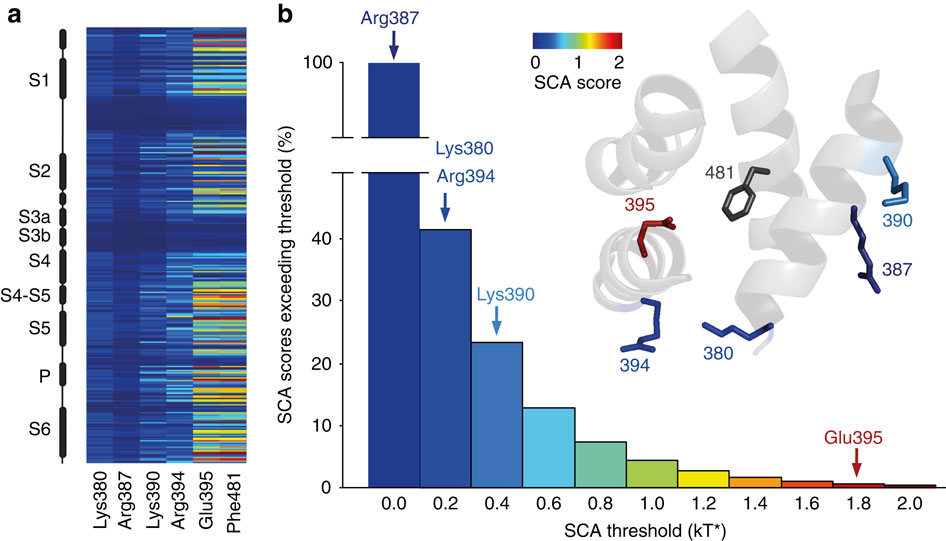
Figure 4: SCA analysis highlights the varying levels of co-evolution between Phe481 and nearby charged side chains. (a) SCA scores of Phe481 and all five charged side chains that are within 12 Å of Phe481 with the vertical axis representing all side chains considered for the SCA analysis (corresponding to positions 208 to 487 inShaker, with black boxes representing α-helical regions in the channel). Scale bar as in (b). (b) Bar diagram indicating the percentage of SCA scores exceeding the binned threshold indicated on the x axis for all pairings in the SCA analysis. The SCA scores of all five charged side chains with respect to Phe481 are highlighted with arrows (Arg387=0.13 kT*, Lys380=0.20 kT*, Arg394=0.21 kT*, Lys390=0.47 kT* and Glu395=1.93 kT*); Note the high co-evolution score for the Glu395-Phe481 coupling. Inset shows the scale of the heat map used in (a,b) and a model displaying all five charged side chains that are within 13 Å of Phe481. Note that the charged side chains are coloured by their SCA score values (Phe481 is shown in dark grey as all SCA scores are relative to Phe481 in this analysis).
Abstract
Voltage-gated potassium channels elicit membrane hyperpolarization through voltage-sensor domains that regulate the conductive status of the pore domain. To better understand the inherent basis for the open-closed equilibrium in these channels, we undertook an atomistic scan using synthetic fluorinated derivatives of aromatic residues previously implicated in the gating of Shaker potassium channels. Here we show that stepwise dispersion of the negative electrostatic surface potential of only one site, Phe481, stabilizes the channel open state. Furthermore, these data suggest that this apparent stabilization is the consequence of the amelioration of an inherently repulsive open-state interaction between the partial negative charge on the face of Phe481 and a highly co-evolved acidic side chain, Glu395, and this interaction is potentially modulated through the Tyr485 hydroxyl. We propose that the intrinsic open-state destabilization via aromatic repulsion represents a new mechanism by which ion channels, and likely other proteins, fine-tune conformational equilibria.
 Model of the pore domain of the tetrameric Kv1.2/2.1 chimera (the voltage-sensing domains are omitted for clarity), with the positions of Phe401, Phe402, Phe481, Phe484 and Tyr485 highlighted in yellow. The inset shows a close-up view of the five aromatic side chains; (b–f) GVs for WT and F3-Phe incorporated at positions 484 (b), 485 (c), 401 (d), 402 (e) and 481 (f); note that at position 485 the native Tyr was also replaced by Phe; insets show currents (−60 to +30 mV in (a,c–f); −60 to +90 mV in (b); all in 10 mV increments) and electrostatic surface potential maps for Phe and F3-Phe (ESP, red=−15 kcal mol−1, green=0 kcal mol−1, blue=+15 kcal mol−1). Scale bar (horizontal), 20 ms; Scale bar (vertical), 5 μA in (c,d–f), 2 μA in (c). WT, V1/2=−23.0±0.5 mV; Phe401TAG+F3-Phe, V1/2=−14.4±0.7; Phe402TAG+F3-Phe, V1/2=−19.0±0.6 mV, Phe484TAG+F3-Phe, V1/2=−24.6±0.6 mV; Tyr485TAG+Phe, V1/2=1.9±5.1 mV, Tyr485TAG+F3-Phe, V1/2=4.6±3.9 mV; Phe481TAG+F3-Phe, V1/2=−45.7±0.8 mV (n=4–10 each). All data=mean±s.e.m.")
 Model showing the tetrameric Kv1.2/2.1 chimera from the intracellular side. The distal part of S6 has been removed for better visibility of Phe481 (Shaker numbering), which is highlighted in yellow. (b) Sequence alignment of the distal end of S6 of various Kv channels: Shaker (GI:288442), Kv1.1 (GI:119395748), Kv2.1 (GI:84570020), Kv3.1 (GI:298603), Kv4.1 (GI:8272404), Kv5.1 (GI:24418476), Kv6.1 (GI:24418479), Kv7.1 (GI:6166005) and Kv8.1 (GI:7657289). The conserved Phe (Phe481 in Shaker) is highlighted in yellow. (c) GVs of Phe and fluorinated Phe derivatives incorporated at position 481 (n=8–10 each; see also Supplementary Table S1); insets show representative currents (−70 to +40 mV in 10 mV increments, F3-Phe reproduced from Fig. 1f) and electrostatic surface potential maps for Phe and F3-Phe (scale as in Fig. 1). (d) ZFV1/2 values plotted against the number of fluorines for Phe and fluorinated Phe derivatives incorporated at position 481. (e) Activation rates plotted against voltage for Phe and fluorinated Phe derivatives incorporated at position 481; insets show sample traces for Phe and F3-Phe (depolarizations in 10 mV increments from a holding potential of -80 mV, traces shown from −70 mV to 40 mV). (f) Deactivation rates plotted against voltage for Phe and fluorinated Phe derivatives incorporated at position 481; insets show sample traces for Phe and F3-Phe (after a depolarizing step to 0 mV different test potentials were applied in 10 mV increments, traces shown from 0 to −100 mV). Scale bar (horizontal), 20 ms in (c,e), Scale bar, 50 ms in (f); Scale bar (vertical), 10 μA in (c,e), Scale bar, 5 μA in (f). All data=mean±s.e.m.")
 Experimentally observed GV data points for Phe and F3-Phe (open and blue circles, respectively; reproduced from Fig. 1f) overlaid by the simulated data based on kinetic modelling using the ZHA model (red lines); (b) Experimentally observed deactivation rates for Phe and F3-Phe (open and blue circles, respectively; reproduced from Fig. 2f) compared with the simulated deactivation rates for Phe and F3-Phe based on kinetic modelling using the ZHA model (red lines); (c) GVs for Phe and F3-Phe introduced on the Ile372Leu, Ser376Thr (LT) double-mutant background (open and blue circles, respectively). The fit for the WT GV is shown in grey, insets show representative currents. Scale bar (horizontal), 20 ms; Scale bar (vertical), 4 μA. (d) Bar diagram comparing the free energy difference between Phe and F3-Phe incorporated at position 481 on the WT (empty bar) or LT double-mutant background (black bar). All experimental data=mean±s.e.m.")
 GVs of Phe and fluorinated Phe derivatives incorporated at position 481 on the background of Lys380Cys (a), Arg387Gln (b), Lys390Gln (c), Arg394Gln (d) and Arg377Lys (e) (n=3–11 each, see also Supplementary Table S1); (f) ΔZFV1/2 values plotted against the number of fluorines for Phe and fluorinated Phe derivatives incorporated at position 481 on the background of Lys380Cys, Arg387Gln, Lys390Gln, Arg394Gln and Arg377Lys. Lines indicate linear fits obtained with f=y0+a*x, with y0=0 (mutants in black, WT in red). All data=mean±s.e.m.")
 Sequence alignment of the S4–S5 linker and the distal part of S6 in various Kv channels (see Fig. 1 for details). The conserved Glu and Phe side chains are highlighted in red and yellow, respectively; other residues tested in this study are highlighted by black boxes (Shaker numbering); (b) Model highlighting the proximity of Glu395 to Phe481 (based on a refined model of the Kv1.2 open channel crystal structure42); (c,d) GVs of Phe and fluorinated Phe derivatives incorporated at position 481 on the background of Glu395Gln (c) and Glu395Asn (d) (n=3–6 each; see also Supplementary Table S1); (e) ΔZFV1/2 values plotted against the number of fluorines for Phe and fluorinated Phe derivatives incorporated at position 481 on the background of Glu395Gln and Glu395Asn (compared with WT). Lines indicate linear fits obtained with f=y0+a*x, with y0=0. All data=mean±s.e.m.")
 and open (lower panel) conformation, respectively. VSD stands for voltage-sensor domain. The close proximity of the Glu395 carboxyl and the Phe481 aromatic moiety in the open state are highlighted by the red oval; the electrostatic repulsion between the two is symbolized by the red arrows. The blue dots represent potassium ions along the permeation pathway. Note that for reasons of simplicity the model depicts an intrasubunit interaction between Glu395 and Phe481, while in the full-length channel the shortest distance between Glu395 and Phe481 is between adjacent subunits, thus indicating an intersubunit interaction.")
