Transport domain unlocking sets the uptake rate of an aspartate transporter
By Nurunisa Akyuz, Elka R. Georgieva, Zhou Zhou, Sebastian Stolzenberg, Michel A. Cuendet, George Khelashvili, Roger B. Altman, Daniel S. Terry, Jack H. Freed, Harel Weinstein, Olga Boudker, and Scott C. Blanchard.
Published in Nature. 2015 Feb 5;518(7537):68-73. PMID: 25652997. PMCID: PMC4351760. [Available on 2015-08-05] Link to publication page.
Projects: The Transport Cycle in Neurotransmitter Uptake Systems, Conformational Dynamics of a Glutamate Transporter Homologue | Core Facility: Computational Modeling
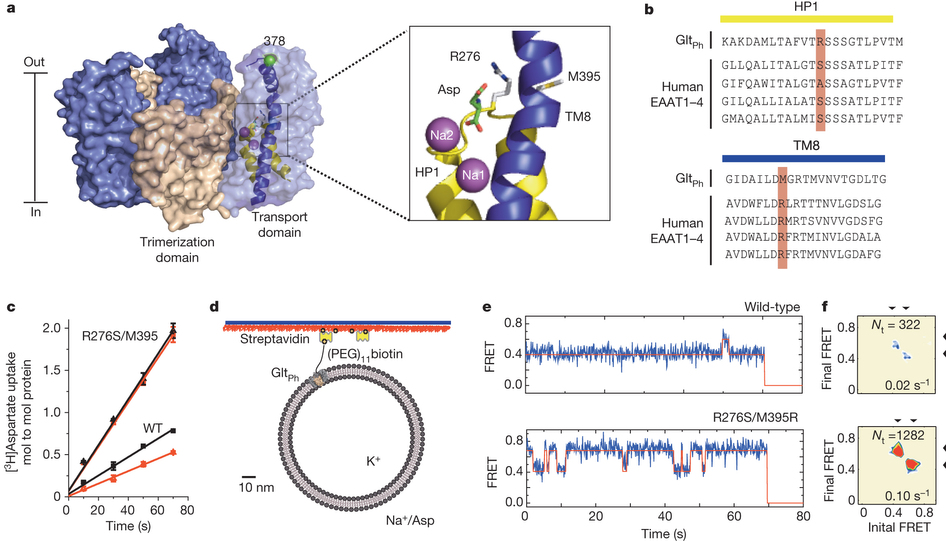
Figure 1. Transport rates and ‘elevator-like’ domain dynamics are correlated. a, Surface representation of the outward-facing GltPh showing the transport (blue) and scaffold (beige) domains. In one protomer, HP1 (yellow) and TM8 (dark blue) are emphasized as cartoons. In the enlarged substrate-binding site (right), mutated residues and aspartate are shown as sticks and ions are shown as spheres. b, Sequence alignment for HP1 and TM8 with mutation sites highlighted in pink. c, Aspartate uptake by unlabelled (black) and labelled (red) transporters. Substrate uptake data are shown as averages of at least three experiments with error bars representing standard deviations. d, Proteoliposome attachment strategy. e, smFRET trajectories recorded for the wild type (WT) and mutant in proteoliposomes under transport conditions. f, Transition density plots corresponding to e. The average transition frequencies and the number of total transitions (Nt) are shown. Colour scale is from tan (lowest) to red (highest frequency).
Abstract
Glutamate transporters terminate neurotransmission by clearing synaptically released glutamate from the extracellular space, allowing repeated rounds of signalling and preventing glutamate-mediated excitotoxicity. Crystallographic studies of a glutamate transporter homologue from the archaeon Pyrococcus horikoshii, GltPh, showed that distinct transport domains translocate substrates into the cytoplasm by moving across the membrane within a central trimerization scaffold. Here we report direct observations of these ‘elevator-like’ transport domain motions in the context of reconstituted proteoliposomes and physiological ion gradients using single-molecule fluorescence resonance energy transfer (smFRET) imaging. We show that GltPh bearing two mutations introduced to impart characteristics of the human transporter exhibits markedly increased transport domain dynamics, which parallels an increased rate of substrate transport, thereby establishing a direct temporal relationship between transport domain motion and substrate uptake. Crystallographic and computational investigations corroborated these findings by revealing that the ‘humanizing’ mutations favour structurally ‘unlocked’ intermediate states in the transport cycle exhibiting increased solvent occupancy at the interface between the transport domain and the trimeric scaffold.
 and cumulative population histograms (right) are shown for the wild type (a) and mutant (c). Experimental conditions are indicated above the panels. Contour plots are colour-coded from tan (lowest) to red (highest population); colour scale from 0–12%. Histograms display the time-averaged state distributions. Solid black lines are fits to sums of individual Gaussian functions (red lines). n is the number of molecules analysed. b, d, Corresponding transition density plots (as in Fig. 1).")
 and higher- (right) FRET states determined for H276,395-GltPh in detergent micelles as functions of Na+ concentration in the presence of 0 (blue), 20 (black) and 100 µM (red) aspartate. Solid lines are fits to Hill equation with Kd = 200, 30 and 15 mM, respectively, and n value of 3. The data points shown are averages and standard errors from three independent biological replicates. b, Logarithmic plots of aspartate Kd values as functions of Na+ concentrations. Data are from isothermal titration calorimetry (ITC) (black) and smFRET (grey). The solid line through the data are a linear fit with slope 3.2. The extent of coupling between Na+ and aspartate binding is similar to wild type (dashed line)23.")
 and average dwell times (b, d) for the low- (low, solid lines) and intermediate- and high-FRET states (int + high, dashed lines) obtained for the wild type (a, b) and H276,395-GltPh (c, d) in detergent. The distributions for apo (blue) and Na+/aspartate-bound proteins (red) were fitted to a probability density function. The fitted time constants are in Extended Data Table 1c. Average dwells are plotted as functions of Na+ concentration in the presence of 10 and 100 µM aspartate for wild type and H276,395-GltPh, respectively. Solid lines are fits to Hill equation with Kd = 15 mM and n = 3.2 for wild type and Kd = 19 mM and n = 3.2 for H276,395-GltPh. The data points shown are averages and standard errors from three independent biological replicates.")
, locked mutant (centre) and unlocked mutant (right) in surface representation, coloured as in Fig. 1, HP2 is red. Residues 276 and 395 are coloured by atom type. The approximate limits of the hydrocarbon layer of the membrane are shown as dashed lines. b, Substrate binding sites (enlarged) viewed from the cytoplasm. HP1 and HP2 are in cartoon representation; aspartate (black) and residues 276, 395 and Asp 394 (coloured by atom type) are emphasized as spheres. Arrowhead (cyan) marks the region of increased solvent accessibility. c, Cytoplasmic view of the unlocked protomer showing the crevice at the domain interface. Dashed line replaces TM2–TM3 loop for clarity. Arrows indicate regions of increased water and lipid accessibility. Open conformations of HP2 were modelled based on the TBOA-bound (green) structure of GltPh3.")
 and H276,395-GltPh (bottom). Periods of long-lived locked states are shaded blue; periods of transitions between unlocked states are shaded red. c, Structures of the inward-facing wild-type locked state (top) and H276,395-GltPh unlocked state (bottom).")
